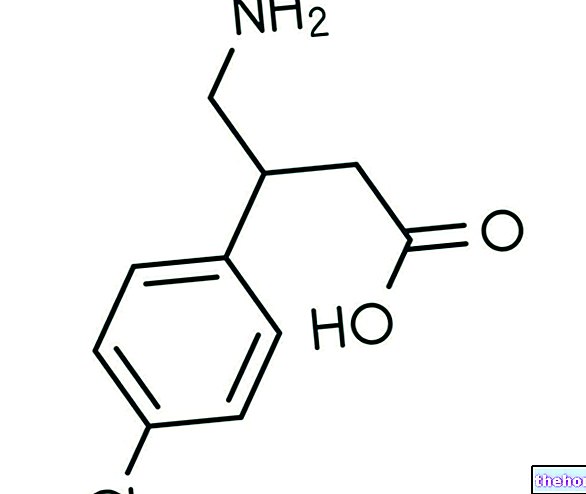
Toimittanut tri Giovanni Chetta
Biokemiasta biomekaniikkaan

Extra-cellular matrix (MEC)
Kuvaus, vaikkakin mitä vähän tiedämme tänään, ECM: stä (solunulkoinen matriisi) on välttämätöntä, jotta ymmärrämme paremmin selkärangan ja asennon muutosten merkityksen terveydelle.
Jokaisen solun, kuten kaikkien monisoluisten elävien organismien, täytyy "tuntea" ja olla vuorovaikutuksessa ympäristönsä kanssa voidakseen suorittaa elintoimintonsa ja selviytyä. Monisoluisessa organismissa solujen on koordinoitava eri käyttäytymismalleja kuin ihmisyhteisössä.

ECM: n kuvataan yleensä koostuvan useista suurista biomolekyyliluokista:
- Rakenneproteiinit (kollageeni ja elastiini)
- Erikoisproteiinit (fibrilliini, fibronektiini, laminiini jne.)
- Proteoglykaanit (aggrekaanit, syndekanit) ja glusaminoglykaanit (hyaluronaanit, kondroitiinisulfaatit, heparaanisulfaatit jne.)
Rakenneproteiineista mm kollageeni muodostavat eläinvaltakunnan eniten edustetun glykoproteiiniperheen. Ne ovat ECM: n läsnäolevia proteiineja (mutta eivät tärkeimpiä) ja ovat varsinaisten sidekudosten (rusto, luu, fascia, jänteet, nivelsiteet) perusaineet.
Kollageenit syntetisoidaan enimmäkseen fibroblasteilla, mutta myös epiteelisolut pystyvät syntetisoimaan niitä.
Kollageenikuidut ovat jatkuvasti vuorovaikutuksessa valtavan määrän muiden ECM -molekyylien kanssa, mikä muodostaa solun elämän kannalta perustavanlaatuisen jatkuvuuden. Liittyvät kuitujen kollageenit ovat hallitsevassa asemassa sellaisten rakenteiden muodostamisessa ja ylläpidossa, jotka kykenevät vastustamaan lähes Elastinen kollageeni tuotetaan ja metaboloituu uudelleen mekaanisen kuormituksen funktiona, ja sen viskoelastisilla ominaisuuksilla on suuri vaikutus ihmisen asentoon.
Kollageenikuiduilla on PG / GAG -pinnoitteensa (proteoglykaanit / glukosaminoglykaanit) ansiosta biosensori- ja biojohtavia ominaisuuksia. Tiedämme itse asiassa, että mikä tahansa mekaaninen voima, joka kykenee synnyttämään rakenteellisen muodonmuutoksen, rasittaa molekyylien välisiä sidoksia, jotka tuottavat pienen sähkövirran, eli pietsosähköisen virran (Athenstaedt, 1969). Siksi kolmiulotteisella ja kaikkialla läsnä olevalla kollageeniverkostolla on myös erityinen ominaisuus biosähköisten signaalien johtamiseen avaruuden kolmessa ulottuvuudessa, joka perustuu kollageenifibrillien ja solujen väliseen suhteelliseen järjestelyyn aferenssisuunnassa (ECM: stä soluihin) tai päinvastoin päinvastoin efferentti.
Kaikki tämä edustaa reaaliaikaista MEC-soluviestintäjärjestelmää ja tällaiset sähkömagneettiset biosignaalit voivat johtaa tärkeisiin biokemiallisiin muutoksiin, esimerkiksi "luun osteoklastit eivät pysty" sulattamaan "pietsosähköisesti varautunutta luuta (Oschman, 2000).
Lopuksi on korostettava, että solu, ei yllättävää, tuottaa jatkuvasti ja huomattavia energiakustannuksia (noin 70%) materiaalia, joka on välttämättä karkotettava yksinomaan varastoimalla protokollageenia (kollageenin biologinen esiaste) tietyissä rakkuloissa ( Albergati, 2004).
Suurin osa selkärankaisista kudoksista vaatii samanaikaisesti kahta elintärkeää ominaisuutta: voimaa ja joustavuutta. Todellinen verkko joustavat kuidut, joka sijaitsee näiden kudosten ECM: n sisällä, sallii palata alkuperäisiin olosuhteisiin voimakkaiden vetojen jälkeen.Elastiset kuidut pystyvät lisäämään elimen tai sen osan venyvyyttä vähintään viisi kertaa. Pitkät, joustamattomat kollageenikuidut ovat välissä joustavien kuitujen välissä, ja niiden tehtävänä on rajoittaa "kudosten vetämisen aiheuttamaa liiallista muodonmuutosta". Elastiini on elastisten kuitujen pääkomponentti ja sitä esiintyy erityisen runsaasti verisuonissa, joilla on joustavat ominaisuudet ( muodostaa yli 50% aortan kuivapainosta), nivelsiteissä, keuhkoissa ja ihossa.
ECM sisältää suuren määrän (eikä vielä tarkasti määriteltyjä) erikoistuneita ei-kollageeniproteiineja, joilla on tyypillisesti spesifisiä sitoutumiskohtia muille ECM-molekyyleille ja solupinnan reseptoreille. Tällä tavalla näiden proteiinien jokainen komponentti toimii "vahvistimena" sekä samanlaisten että erilaisten molekyylien välisissä yhteyksissä ja luo äärettömän biokemiallisen verkon, joka kykenee tuottamaan, muuntamaan, vaihtamaan ja lisäämään jopa etäisyydellä miljoonia ja miljoonia biokemiallisia tietoja (ja energiaa).
Tärkeä "solunulkoisen matriisin erikoistunut proteiini on fibronektiini, korkean molekyylipainon glykoproteiini, jota löytyy kaikista selkärankaisista. Fibronektiini näyttää kykenevän vaikuttamaan solujen kasvuun, solujen väliseen tarttumiseen ja ECM: llä, solujen siirtymiseen eri tavoin (solu voi liikkua jopa 5 cm päivässä - Albergati, 2004) jne. Tunnetuin isoformi, tyyppi III, sitoutuu integriineihin . Jälkimmäiset ovat transmembraaniproteiinien perhe, jotka toimivat mekaanoreseptoreina: ne antavat valikoivasti ja moduloitavalla tavalla mekaanisia vetoja ja työntöjä ECM: stä solun sisällä ja päinvastoin, aiheuttaen sytoplasmassa useita reaktioita, joihin liittyy sytoskeletti ja muita proteiineja, jotka säätelevät solujen tarttumista, kasvua ja migraatiota (Hynes, 2002).
Glukosaminoglykaanit (GAGS) ja proteoglykaanit (PG) muodostavat erittäin hydratoidun geelimäisen aineen, joka on määritelty sidekudoksissa ja jossa fibrillaariset proteiinit sijaitsevat ja imeytyvät. Tämä polysakkaridigeelin muoto kykenee toisaalta sallimaan ECM: n kestämään huomattavia puristusvoimia ja toisaalta sallimaan ravinteiden, metaboliittien ja hormonien nopean, jatkuvan ja valikoivan diffuusion veren ja kudosten välillä.
Glukosaminoglykaanien polysakkaridiketjut ovat volumetrisesti liian jäykkiä taittuakseen polypeptidiketjuille tyypillisten pallomaisten rakenteiden sisään, lisäksi ne ovat erittäin hydrofiilisiä. ne vievät suuren määrän suhteessa niiden massaan ja muodostavat siten huomattavia määriä geeliä myös pieninä pitoisuuksina. Näistä hallitseva rooli on Na +: lla, joka antaa koko osmoottisen kapasiteetin ja vangitsee valtavan määrän vettä ECM: ään. Tällä tavoin syntyy turvotuksia (turgoreita), joiden avulla ECM voi vastustaa jopa tärkeitä puristusvoimia (tämän ansiosta esimerkiksi lonkan rusto voi fysiologisissa olosuhteissa kestää täydellisesti useiden satojen ilmakehän paineen).
Sidekudoksen sisällä GAG: t edustavat alle 10-12% kokonaispainosta, mutta ominaisuuksiensa vuoksi ne täyttävät monia solunulkoisia tiloja muodostaen erikokoisia ja -tiheyksisiä sähkövarauksia sisältävän hydratoidun geelin huokosia, jotka toimivat siten valikoivina avainpisteitä tai "palvelimia", joiden kautta MEC: n sisällä olevien molekyylien ja solujen liikennettä säännellään niiden koon, painon ja sähkövarauksen perusteella.
Hyaluronihappo (hyaluronaani, hyaluronaatti) edustaa ehkä yksinkertaisinta GAG: sta. Kokeelliset ja molekyylibiologiset tiedot vahvistavat, että sillä on keskeinen rooli luiden ja nivelten tasolla kestävyydessä huomattavia paineita vastaan. : se luo tyhjiä tiloja solujen väliin, joihin ne siirtyvät myöhemmässä vaiheessa (Albergati, 2004).
ECM ei eritä kaikkia PG: itä, jotkut ovat plasmakalvojen kiinteitä komponentteja (Alberts, 2002).
Solunulkoista matriisia voidaan siksi pitää erittäin monimutkaisena verkostona, jossa proteiinit, PGS ja GAG tarjoavat lukemattomia toimintoja, mukaan lukien rakennetuki ja jokaisen kudoksen ja orgaanisen toiminnan säätely. Globaalia solun homeostaasia on pidettävä mekanismikompleksina, joka voi syntyä ja kehittyä solun sisällä tai ECM: n ulkopuolella; jälkimmäisessä tapauksessa solu voi edustaa väli- tai lopullista kohdetta. Sen lisäksi, että solunulkoiset komponentit edustavat solurakenteiden fyysisiä tukirakenteita, ne toimivat myös todellisina kohteina elintärkeiden prosessien käynnistämiselle, kehittämiselle ja lopettamiselle sekä endosellulaarisessa ympäristössä että elimissä ja järjestelmissä. Edessämme on ääretön biokemiallinen verkko, joka kykenee tuottamaan, muuntamaan, vaihtamaan ja lisäämään miljoonien ja miljoonien tietojen määrää jopa etäisyyden päästä.
Jokainen kehon solu on jatkuvasti vuorovaikutuksessa ECM: n kanssa sekä mekaanisten että kemiallisten ja energeettisten näkökohtien suhteen, ja sillä on "dramaattisia" vaikutuksia kudosten staattiseen ja dynaamiseen arkkitehtuuriin. P. A. Baccin mukaan interstitiaalinen matriisi edustaa todella elintärkeiden reaktioiden äitiä, paikkaa, jossa ennen kaikkea aineen ja energian vaihto tapahtuu. Kaikki kudokset on kytketty toisiinsa ja toiminnallisesti integroitu toisiinsa ei suljetuissa mutta avoimissa järjestelmissä; niiden välillä tapahtuu jatkuvaa vaihtoa, joka voi tapahtua sekä paikallisesti että systeemisesti hyödyntäen biokemiallisia, biofysikaalisia ja sähkömagneettisia viestejä eli käyttämällä erilaisia energiamuotoja.
Kuten F. G.Majoitettuna solu ja solunulkoinen matriisi edustavat kahta vain näennäisesti erillistä maailmaa, joiden on välttämättä koko elämän ajan oltava vuorovaikutuksessa milloin tahansa toimiakseen oikein ja synergistisesti. Tämä vaatii poikkeuksellisen sarjan signaaleja, joita seuraa yhtä uskomaton molekyylibiologisten toimintojen sarja.
Muut artikkelit aiheesta "Extra -Cellular Matrix - rakenne ja toiminnot"
- Skolioosin hoito
- Skolioosi - syyt ja seuraukset
- Skolioosin diagnoosi
- Skolioosin ennuste
- Sidekudos ja sidekudos
- Liitäntänauha - ominaisuudet ja toiminnot
- Asento ja jännitys
- Ihmisen liike ja ratsastustuen merkitys
- Oikeiden ratsastus- ja okklusaalituen merkitys
- Idiopaattinen skolioosi - myyttejä hävitettäväksi
- Skolioosin kliininen tapaus ja terapeuttinen protokolla
- Hoidon tulokset Kliininen tapaus skolioosi
- Skolioosi luonnollisena asenteena - Bibliografia