Jotta voisimme puhua ymmärrettävästi "hemoglobiini (Hb), on hyödyllistä hoitaa ensimmäinen myoglobiini (Mb) joka on hyvin samanlainen kuin hemoglobiini, mutta on paljon yksinkertaisempi. heme.
Myoglobiini on pallomainen proteiini, joka koostuu yhdestä noin viisikymmentä aminohappoa sisältävästä ketjusta (se riippuu organismista) ja sen molekyylipaino on noin 18 Kd.
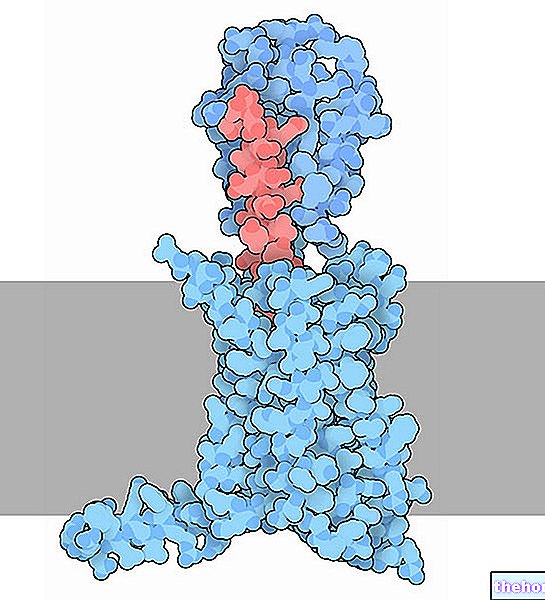
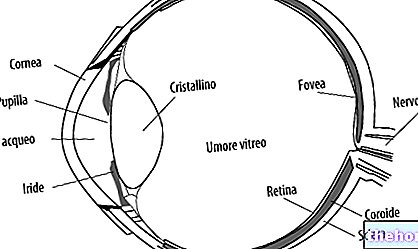
Kuten mainittiin, se on varustettu hemiryhmällä, joka on insertoitu proteiinin hydrofobiseen (tai lipofiiliseen) osaan, joka koostuu taitteista, jotka johtuvat kuituproteiinien a-helix-rakenteista.
Myoglobiini koostuu pääasiassa a-heliksien segmenteistä, joita on kahdeksan, ja se koostuu lähes yksinomaan ei-polaarisista tähteistä (leusiini, valiini, metioniini ja fenyylialaniini), kun taas polaarisia tähteitä ei käytännössä ole (asparagiinihappo, glutamiinihappo, lysiini) ja arginiini); ainoat polaariset tähteet ovat kaksi histidiiniä, joilla on keskeinen rooli hapen sitoutumisessa hemiryhmään.
Hemiryhmä on kromoforiryhmä (imeytyy näkyvään) ja on myoglobiinin funktionaalinen ryhmä.
Katso myös: glykoitunut hemoglobiini - hemoglobiini virtsassa
Vähän kemiaa

Protoporfyriinin ja raudan välinen sidos on tyypillinen koordinaatioyhdisteiden sidos, jotka ovat kemiallisia yhdisteitä, joissa keskiatomi (tai ioni) muodostaa sidoksia muiden kemiallisten lajien kanssa suuremmalla määrällä kuin sen hapetusluku (sähkövaraus). Hemen tapauksessa nämä sidokset ovat palautuvia ja heikkoja.
Raudan koordinaatioluku (koordinaatiosidosten lukumäärä) on kuusi: raudan ympärillä voi olla kuusi molekyyliä, jotka jakavat sidoselektronit.
Koordinointiyhdisteen muodostamiseksi tarvitaan kaksi kiertoradaa, joilla on oikea suunta: toinen kykenee "hankkimaan" elektroneja ja toinen lahjoittamaan niitä.
Hemessä rauta muodostaa neljä tasomaista sidosta, joissa on neljä typpiatomia proto-porfyriinirenkaan keskellä ja viides sidos proksimaalisen histidiinitypen kanssa; raudalla on kuudes vapaa koordinointiside ja se voi sitoutua happiin.
Kun rauta on vapaan ionin muodossa, sen tyyppi kiertää d heillä kaikilla on sama energia; myoglobiinissa rauta -ioni sitoutuu protoporfyriiniin ja histidiiniin: nämä lajit häiritsevät magneettisesti kiertoradia d rautaa; häiriön laajuus on erilainen eri kiertoradilla d riippuen niiden alueellisesta suuntauksesta ja häiritsevistä lajeista. Koska orbitaalien kokonaisenergian on oltava vakio, häiriö aiheuttaa energeettisen erottelun eri orbitaalien välillä: joidenkin orbitaalien hankkima energia vastaa muiden menettämää energiaa.
Jos orbitaalien välinen etäisyys ei ole kovin suuri, on suositeltavaa käyttää suurta spin-elektronista järjestelyä: sitovat elektronit pyrkivät järjestäytymään rinnakkaiskierroksiin mahdollisimman monella alatasolla (suurin moninkertaisuus); jos toisaalta häiriö on erittäin voimakasta ja orbitaalien välillä on suuri ero, voi olla helpompaa yhdistää sidoselektronit alemman energian (matala spin) orbitaaleille.
Kun rauta sitoutuu happiin, molekyyli olettaa matalan spin -järjestelyn, kun taas raudalla on kuudes koordinaatiosidos vapaa, molekyylillä on korkea spin -järjestely.
Tämän spin -eron ansiosta pystymme ymmärtämään myoglobiinin spektrianalyysin avulla, onko happi (MbO2) sitoutunut siihen vai ei (Mb).
Myoglobiini on tyypillinen lihasproteiini (mutta sitä ei löydy vain lihaksista).
Myoglobiini uutetaan siittiövalasta, jossa sitä on suuria määriä, ja puhdistetaan sitten.
Valaiden hengitys on samanlainen kuin ihmisillä: keuhkoineen niiden on imettävä ilmaa hengitysprosessin kautta; siittiöiden on tuoda mahdollisimman paljon happea lihaksiin, jotka kykenevät keräämään happea sitomalla sen niissä olevaan myoglobiiniin; happi vapautuu sitten hitaasti valaan upotettaessa, koska sen aineenvaihdunta vaatii happea: mitä suurempi määrä happea, jonka siittiövalas pystyy absorboimaan ja mitä enemmän happea on saatavilla sukelluksen aikana.
Myoglybiini sitoo happea palautuvalla tavalla ja on läsnä perifeerisissä kudoksissa suuremmassa osassa sitä enemmän, mitä enemmän kudos on tottunut työskentelemään ajassa kaukana olevien hapensaantien kanssa.
<--- Myoglobiini on lihaksissa oleva proteiini, jonka tehtävä on juuri hapen "säiliö".

Se, mikä tekee lihan enemmän tai vähemmän punaiseksi, on hemoproteiinien pitoisuus (heemi tekee lihan punaiseksi).
Hemoglobiinilla on monia rakenteellisia yhtäläisyyksiä myoglobiinin kanssa ja se pystyy sitomaan molekyylipitoista happea palautuvalla tavalla; mutta vaikka myoglobiini rajoittuu lihaksiin ja perifeerisiin kudoksiin yleensä, hemoglobiinia löytyy punasoluista tai punasoluista (ne ovat pseudosoluja eli ne eivät ole todellisia soluja), jotka muodostavat 40% verestä.
Toisin kuin myoglobiini, hemoglobiinin tehtävä on ottaa happea keuhkoihin, vapauttaa se soluihin, missä sitä tarvitaan, ottaa hiilidioksidi ja vapauttaa se keuhkoihin, joissa sykli alkaa uudelleen.
L "hemoglobiini se on tetrametri, eli se koostuu neljästä polypeptidiketjusta, joissa kussakin on hemiryhmä ja jotka ovat identtiset kaksittain (ihmisessä on kaksi alfa- ja kaksi beeta -ketjua).
Hemoglobiinin päätehtävä on hapen kuljettaminen; toinen veren tehtävä, johon hemoglobiini liittyy, on aineiden kuljettaminen kudoksiin.
Polulla keuhkoista (runsaasti happea) kudoksiin hemoglobiini kuljettaa happea (samalla kun muut aineet pääsevät kudoksiin), kun taas päinvastoin se kuljettaa mukanaan kudosten keräämää jätettä, erityisesti hiiltä aineenvaihdunnassa syntyvää dioksidia.
Ihmisen kehityksessä on geenejä, jotka ilmentyvät vain tietyn ajan; tästä syystä on olemassa erilaisia hemoglobineja: sikiön, alkion, aikuisen miehen.
Näiden eri hemoglobiinien muodostavilla ketjuilla on erilaiset rakenteet, mutta joillakin samankaltaisuuksilla itse asiassa niiden suorittama toiminto on suunnilleen sama.
Selitys useiden eri ketjujen olemassaololle on seuraava: organismien evoluutioprosessin aikana jopa hemoglobiini on kehittynyt erikoistuneena hapen kuljettamiseen rikkailta alueilta puutteellisille alueille. " evoluutioketjun l "hemoglobiini kuljetti happea pienissä organismeissa; evoluution aikana organismit saavuttivat suurempia ulottuvuuksia, joten hemoglobiinia muutettiin siten, että se pystyi kuljettamaan happea kauempana oleville alueille, joissa se oli runsaasti sitä; tätä varten ne on koodattu evoluutioprosessin aikana hemoglobiinin muodostavien ketjujen uusiksi rakenteiksi.
Myoglobiini sitoo happea jopa vaatimattomissa paineissa; perifeerisissä kudoksissa paine (PO2) on noin 30 mmHg: myoglobiini ei vapauta happea tässä paineessa, joten se olisi tehoton hapen kantajana. , sillä on joustavampi käyttäytyminen: se sitoo happea korkeisiin paineisiin ja vapauttaa sen, kun paine laskee.
Kun proteiini on toiminnallisesti aktiivinen, se voi muuttaa muotoaan hieman; esimerkiksi happipitoisella myoglobiinilla on erilainen muoto kuin hapettomalla myoglobiinilla, eikä tämä mutaatio vaikuta sen naapureihin.
Tilanne on erilainen assosioituneiden proteiinien, kuten hemoglobiinin, tapauksessa: kun ketju hapettaa, sen indusoidaan muuttavan muotoaan, mutta tämä muutos on kolmiulotteinen, joten myös muut tetrametrin ketjut vaikuttavat siihen. ehdottaa, että yhden muutos vaikuttaa muihin naapureihin, vaikkakin eri tavalla; kun ketju hapettaa, muut tetrametrin ketjut ottavat "vähemmän vihamielisen asenteen" happea kohtaan: ketjun vaikeus hapettuminen vähenee, kun sen lähellä olevat ketjut hapettavat vuorotellen. Sama koskee hapenpoistoa.
Deoksihemoglobiinin kvaternaarista rakennetta kutsutaan T (jännittynyt) muotoksi, kun taas oksihemoglobiinin rakennetta kutsutaan R (vapautuneeksi) muotoksi; jännittyneessä tilassa happamien aminohappojen ja emäksisten aminohappojen välillä on sarja melko voimakkaita sähköstaattisia vuorovaikutuksia, jotka johtavat deoksiehemoglobiinin jäykään rakenteeseen (tästä syystä "jännittynyt muoto"), kun taas happi on kytketty, näiden kokonaisuus vuorovaikutus vähenee (tästä syystä "vapautettu muoto"). Lisäksi hapen puuttuessa histidiinin varaus (katso rakenne) stabiloituu asparagiinihapon vastaisella varauksella, kun taas hapen läsnä ollessa proteiinilla on taipumus menettää protoni; Kaikki tämä liittyy siihen, että hapetettu hemoglobiini on vahvempi happo kuin hapetettu hemoglobiini: bohrin vaikutus.
PH -arvosta riippuen hemiryhmä sitoutuu enemmän tai vähemmän helposti happiin: happamassa ympäristössä hemoglobiini vapauttaa happea helpommin (jännittynyt muoto on vakaa), kun taas emäksisessä ympäristössä sidos hapen kanssa on vaikeampi.

Jokainen hemoglobiini vapauttaa 0,7 protonia per happea (O2).
Bohrin vaikutuksen ansiosta hemoglobiini voi parantaa kykyä kuljettaa happea.
Hemoglobiinin, joka kulkee keuhkoista kudoksiin, on tasapainotettava paineen, pH: n ja lämpötilan funktiona.
Katsotaanpa lämpötilan vaikutusta.
Keuhkoalveolien lämpötila on noin 1-1,5 ° C alhaisempi kuin ulkolämpötila, kun taas lihaksissa lämpötila on noin 36,5-37 ° C; kun lämpötila nousee, kyllästyskerroin laskee (samalla paineella): tämä tapahtuu, koska liike -energia kasvaa ja dissosiaatio suosii.

On muitakin tekijöitä, jotka voivat vaikuttaa hemoglobiinin kykyyn sitoutua happiin, joista yksi on 2,3 -bisfosfoglysereraatin pitoisuus.
2,3-bisfosfoglysereraatti on aineenvaihdunta, jota esiintyy punasoluissa 4-5 mM pitoisuutena (missään muussa kehon osassa sitä ei ole näin suurena pitoisuutena).
Fysiologisessa pH: ssa 2,3 -bisfosfogloseraatti deprotonoidaan ja siinä on viisi negatiivista varausta; se on kiinnitetty kahden hemoglobiinin beeta -ketjun väliin, koska näillä ketjuilla on korkea positiivisten varausten pitoisuus. Beetaketjujen ja 2,3 -bisfosfoglysereraatin väliset sähköstaattiset vuorovaikutukset antavat järjestelmälle tietyn jäykkyyden: saadaan jännittynyt rakenne, jolla ei ole juurikaan affiniteettia hapelle; hapetuksen aikana 2,3 -bisfosfoglysereraatti karkottaa.
Erytrosyytteissä c "on erityinen laite, joka muuntaa 1,3-bisfosfogloseraatin (aineenvaihdunnan tuottama) 2,3-bisfosfoglysereraatiksi siten, että sen pitoisuus on 4-5 mM ja siksi hemoglobiini pystyy vaihtamaan" happea "kudoksissa.
Kudokseen saapuva hemoglobiini on vapautetussa tilassa (sitoutunut happiin), mutta kudoksen läheisyydessä se on karboksyloitu ja siirtyy jännittyneeseen tilaan: tässä tilassa oleva proteiini on vähemmän taipuvainen sitoutumaan hapen kanssa vapautettuun tilaan, siksi hemoglobiini vapauttaa happea kudokseen; Lisäksi veden ja hiilidioksidin välisessä reaktiossa muodostuu H + -ioneja ja siten lisää happea bohrin vaikutuksesta.
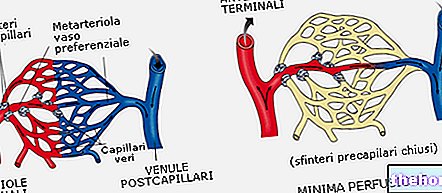
Hiilidioksidi diffundoituu erytrosyyttiin, joka kulkee plasmakalvon läpi; Koska punasolut muodostavat noin 40% verestä, meidän pitäisi odottaa, että vain 40% kudoksista leviävästä hiilidioksidista pääsee niihin, itse asiassa 90% hiilidioksidista pääsee punasoluihin, koska ne sisältävät entsyymiä, joka muuttaa hiilidioksidia hiilihapossa se johtaa siihen, että hiilidioksidin kiinteä pitoisuus punasoluissa on alhainen ja siksi sisäänpääsynopeus on korkea.

Toinen ilmiö, joka ilmenee, kun erytrosyytti saavuttaa kudoksen, on seuraava: gradientin mukaan "HCO3- (hiilidioksidijohdannainen) poistuu" erytrosyytistä ja negatiivisen varauksen tuoton tasapainottamiseksi meillä on "klorideja, jotka määrittää osmoottisen paineen nousun: tämän vaihtelun tasapainottamiseksi on myös vettä, joka aiheuttaa punasolujen turvotusta (HAMBURGER -vaikutus). Päinvastainen ilmiö ilmenee, kun erytrosyytti saavuttaa keuhkoalveolit: punasolujen deflaatio (HALDANE -vaikutus) Siksi laskimopunasolut (suunnattu keuhkoihin) ovat pyöreämpiä kuin valtimot.