«Glykolyysin ensimmäiset vaiheet
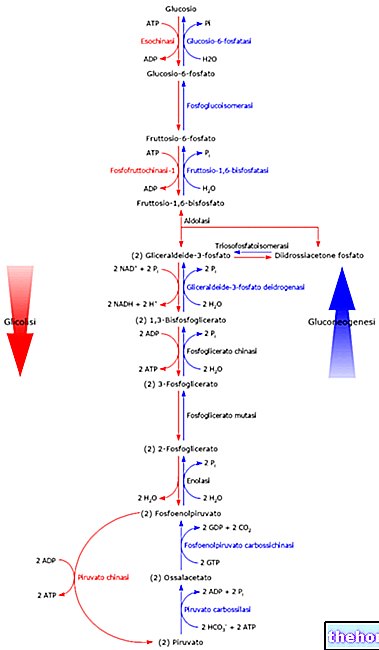
Ensimmäinen glykolyysin toisessa vaiheessa käytetty entsyymi on glyserraldehydi-3-fosfaattidehydrogenaasi; dehydrogenaasit ovat entsyymejä, jotka katalysoivat pelkistävän voiman siirtymistä pelkistävästä molekyylistä, joka hapettuu toiseen pelkistyneeseen molekyyliin (redoksireaktio) .Ensyymin substraatit ovat NAD (nikotidamidiadeniinidinukleotidi) ja FAD (flaviiniadeniinidinukleotidi).
Tässä vaiheessa dehydrogenaasi katalysoi glyserraldehydi-3-fosfaatin muuttumisen 1,3-bisfosfogloseraatiksi: samalla katalyyttikohdalla aldehydiryhmä hapetetaan karboksyyliksi, minkä seurauksena NAD + pelkistyy NADH: ksi, ja sen jälkeen karboksyyliryhmä pystyy anhydridisidoksen muodostamiseksi ortofosfaatin kanssa. Ensimmäinen prosessi on erittäin eksergoninen (se vapauttaa energiaa), kun taas toinen on erittäin eksergoninen (se vaatii energiaa); jos katalyyttipaikkaa ei olisi, globaalia reaktiota ei tapahtuisi: ensimmäinen reaktio tapahtuisi, kun vapautuisi energiaa, joka dispergoituisi lämmöksi ja jota ei siten voida käyttää muodostamaan anhydridisidos.

1,3-bisfosfogloseraatin muodostumisen jälkeen entsyymi palauttaa lähtörakenteensa ja on valmis vaikuttamaan uuteen substraattiin.

Sitten tulee fosfoglysereraattikinaasi joka mahdollistaa fosforyylin siirtämisen 1,3-bisfosfoglysereraatista ADP-molekyyliin; olemme saaneet ATP: tä (yksi ATP kutakin glyseraldehydi-3-fosfaattimolekyyliä kohden, siis kaksi ATP: tä kutakin glukoosimolekyyliä kohti), joka kompensoi glykolyysin ensimmäinen vaihe.
Arsenaattianioni (AsO43-) vaikuttaa glykolyyttiseen reittiin, koska se voi korvata fosfaatin glykolyysin toisen vaiheen ensimmäisessä reaktiossa, jolloin saadaan 1-arsenio-3-fosfoglysereraattia, joka on erittäin epävakaa ja heti kun se on vapaa katalyytistä hydrolysoi vapauttaen "arsenaatin, joka palaa kiertoon. Siksi arsenaatti jäljittelee fosfaatin vaikutusta ja siirtyy katalyyttikohtaan: arsenaatin läsnä ollessa ATP: tä tuottavaa reaktiota (1,3-bisfosfoglysereraatista 3-fosfoglysereraatiksi) ei tapahdu, koska 3-fosfaattiglysereraldehydi muunnetaan suoraan 3-fosfoglysereraatiksi; ilman ATP: tä solut kuolevat (arseenihappomyrkytys).
Oksidatiivisen vaiheen kolmannessa reaktiossa 3-fosfoglysereraatti muuttuu 2-fosfoglysereraatiksi fosfoglysereraattimutaasi; reaktio käsittää 2,3-bisfosfoglysereraattivälituotteen.

Seuraavassa vaiheessa entsyymi puuttuu asiaan enolaasi joka katalysoi vesimolekyylin poistumista 2-fosfoglysereraatin hiilipitoisesta rungosta, jolloin saadaan pyrivoitu fosfenoli (PEP);

PEP: llä on suuri potentiaali siirtää fosforyyliä: se siirtyy entsyymin vaikutuksesta pyruvaatikinaasi, fosforyyli ADP: ksi, jolloin saadaan ATP: tä toisen vaiheen viidennessä vaiheessa, jolloin saadaan pyruvaattia.

2-fosfoglysereraatilla ja 3-fosfoglysereraatilla on alhainen fosforyylin siirtoteho, joten ATP: n saamiseksi näistä molekyyleistä 3-fosfoglysereraatti muutetaan glykolyysin aikana 2-fosfoglysereraatiksi, koska se saadaan jälkimmäisestä PEP: stä, joka on lajeja, joilla on suuri siirtymismahdollisuus.
Ennen kuin jatkamme, avataan sulkeet 2,3-bisfosfoglysereraatista; jälkimmäinen on läsnä kaikissa soluissa, joissa glykolyysi tapahtuu hyvin pieninä pitoisuuksina (se on glykolyysin toisen vaiheen kolmannen reaktion välituote). Toisaalta punasoluissa 2,3-bisfosfogloseraatin kiinteä pitoisuus on 4-5 mM (maksimikonsentraatio), koska niillä on entsymaattinen perintö, jonka tehtävänä on tuottaa se; punasoluissa poikkeama glykolyysistä tuottaa 2,3-bisfosfoglysereraattia: 1,3-bisfosfoglysereraatti muuttuu 2,3-bisfosfoglysereraatiksi bisfosfoglysereraattimutaasi (erytrosyytti) ja 2,3-bisfosfogloseraatti bisfosfoglysereraattifosfataasi (erytrosyytti) muuttuu 3-fosfoglysereraatiksi. Sitten erytrosyyteissä osa glykolyysistä saadusta 1,3-bisfosfoglysereraatista muutetaan 2,3-bisfosfoglysereraatiksi, joka palaa sitten glykolyyttiseen reittiin 3-fosfoglysereraattina; tällöin hapetusvaiheen kolmas vaihe menetetty ATP-määrä on hinta, jonka erytrosyytti on valmis maksamaan pitääkseen yllä näiden solujen tarvitseman 2,3-bisfosfoglysereraatin pitoisuuden, koska se vaikuttaa "hemoglobiinin sitomiskykyyn" happea.
Olemme nähneet, että glykolyysin toisen vaiheen ensimmäisessä reaktiossa NAD + pelkistyy NADH: ksi, mutta on välttämätöntä, että pyruvaatin saamisen jälkeen NADH muutetaan NAD +: ksi: tämä tapahtuu maitohappokäymisen yhteydessä (saadaan laktaattia) tai alkoholikäymisellä (pyruvaatidekarboksylaasi, joka dekarboksyloi pyruvaatin ja dehydrogenaasi, joka muodostaa etanolin) tulevat peliin; käyminen ei sisällä happea (anaerobit).
Maitohappokäymisen vuoksi maitohappo, jos sitä ei hävitetä riittävästi, kerääntyy lihaksiin ja vapauttaa H +: ta, aiheuttaa lihasten tahatonta supistumista ja siten kouristuksia; voimakkaan stressin lihaksen pH voi olla vähintään 6,8.
Cori -syklin kautta osa lihaksen väsymyksestä siirtyy maksaan, kun lihas on ylikuormitettu. Oletetaan, että lihas toimii ilman happea (väärä oletus): jos lihas toimii kohtalaisesti, supistumiseen tarvittava ATP saadaan yksinomaan glykolyysin avulla. Jos lihaksen aktiivisuus lisääntyy ja tarvitaan lisää ATP: tä, nopeuta aerobista aineenvaihduntaa laktaatti, joka siten hävitetään glukoosiksi. Todellisuudessa lihas hyödyntää aerobista aineenvaihduntaa: jos happea on saatavilla, lihas hyödyntää ennen kaikkea aerobisen aineenvaihdunnan tarjoamaa ATP: tä ja kun happea ei ole enää saatavilla, anaerobinen aineenvaihdunta kiihtyy Cori -syklin kautta Tässä syklissä oletetaan, että laktaatti siirtyy lihasta maksaan, jossa energiaa kuluttamalla syntyy enemmän glukoosia, joka palaa lihakseen. Tämän syklin aikana osa lihaksessa kulutetusta ATP: stä saadaan maksasta, glukoneogeneesiprosessin kautta pystyy tuottamaan glukoosia, jota lihakset voivat käyttää ATP: n saamiseen.
Tähän mennessä kuvattu glukoosin aineenvaihdunta ei sisällä happea, mutta glukoosin aerobinen metabolia mahdollistaa 17-18 kertaa suuremmat määrät ATP: tä kuin glykolyyttisen reitin, joten kun solulla on mahdollisuus valita aerobisen ja ed. anaerobe, suosii ensimmäistä.
Aerobisessa aineenvaihdunnassa pyruvaatti tulee mitokondrioihin, missä se muuttuu ja lopulta saadaan hiilidioksidia ja vettä; tällä tavalla saadaan 34 ATP -molekyyliä kullekin hajonneen glukoosin molekyylille.