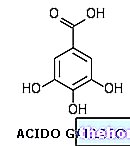
Skikimiinihapporeitti: toissijainen metaboliareitti, jonka esiasteena on skikimihappo, molekyyli, joka sisältää itsessään ne rakenteelliset ja kemialliset ominaisuudet, jotka löytyvät siitä peräisin olevista toissijaisista metaboliiteista.
Skichimiinihappomolekyyli koostuu: 6-jäsenisestä renkaasta, 1 karboksyyliryhmästä ja 3 hydroksyyliryhmästä. Sama molekyylirakenne löytyy siitä peräisin olevista sekundaarisista metaboliiteista, joita itse asiassa kutsutaan happo, skitsimikko. Skikimiinihappo on peräisin kahden välituotteen yhdistämisestä kahdella eri primaarisella metaboliareitillä:
erytrosium-4-fosfaatti (3C): fotosynteesin pimeän vaiheen välituote, anabolinen aineenvaihduntaprosessi;
fosfenolipyruviinihappo (3C): glykolyysin välituote, katabolinen metabolinen prosessi;
Siksi erytrosium-4-fosfaatti + fosfenolipyruviinihappo = skitsimiinihappo: sekundaaristen metaboliareittien ensimmäinen edeltäjä.
Solu syntetisoi skitsimiinihapon silloin, kun tarpeet sen sallivat, tai kun kahden ensisijaisen välituotteen määrät ovat niin suuria ja sellaisia, että ne voivat kerääntyä; tämä tapahtuu, kun solussa on riittävä määrä ATP: tä hidastaa primaarisen katabolian ja anabolian reaktioita.
Maloni- ja mevalonihapon polku: molemmat esiasteet ovat peräisin asetyyli -CoA -molekyylistä, joten molemmat ovat yhdellä polulla: asetaattireitti. solun ensisijainen aineenvaihdunta.
Asetaattiryhmä (ryhmä, jossa on kaksi hiiliatomia) + CoA (koentsyymi A) = asetyyli -CoA: ensisijaiseen aineenvaihduntaan kuuluva molekyyli, jota käytetään biologisena rakennuspalikkana sekundaaristen metaboliittien rakentamisessa.
Asetaattireitti erotetaan sitten malonihapporeittiä ja mevalonihapporeittiä käyttäen.Koentsyymi A toimii kahden hiilipitoisen yksikön kuljetuksena sytoplasmasta solun mitokondrioon, jossa Krebs -sykli tapahtuu. Hiili kuljetetaan sen sijaan muualle energiaylijäämän sattuessa ja muodostaen monipuolisimmat toissijaiset metaboliitit; näillä on parillinen määrä hiiliatomeja, kuten malonihappo (C4) ja mevaloni (C6).
Siksi skitsimiinihapon ja asetaatin metabolisilla reiteillä on tarkka molekyylirakenne, jonka avulla voimme helposti tunnistaa niiden toissijaiset johdannaiset. Varten the alkaloidit, joilla on monipuolinen arkkitehtuuri, esiasteen tunnistaminen ei ole niin helppoa; toisin sanoen ei ole niin helppoa luokitella yksittäisiä alkaloidiluokkia jäljittämällä jokainen yksittäinen esiaste. Alkaloideilla on itse asiassa enemmän kuin yksi esiaste, koska ne ovat peräisin aminohapoista (primaarisista typpiyhdisteistä, joita solu käyttää sekundääristen typpimolekyylien tuottamiseen). Toissijaiset typpimetaboliitit ovat pääasiassa alkaloideja, mutta on myös muita molekyylejä, joiden terveysprofiili on heikko glykosidit (katkerat mantelit) ja β-syaanit (pigmentit) Aminohapot ovat typpipitoisia yhdisteitä, jotka ovat monipuolistuneet toisistaan, ja tämä monimuotoisuus heijastaa niiden suorien johdannaisten, jotka ovat alkaloideja, monipuolistumista.
Ainoa kemiallinen elementti, joka yhdistää eri alkaloidiluokat, on typpiatomi, joka on suljettu heterosykliseen renkaaseen, tai ainakin typpiatomi, jossa on vapaa elektronidubletti, joka antaa niille perusominaisuuksia; sama perusreaktiivisuus, jonka avulla voimme erottaa yksittäiset alkaloidit siirtymän kautta.
Voimme tiivistää sen sanomalla hiilihydraattireitti on metabolinen reitti, joka on kaikkien toissijaisten metaboliittien synteesin perustasiksi se sisältää kaikki aiemmin havaitut metaboliset reitit:
- asetaatti on glukoosimolekyylin täydellisen purkamisen tuote;
- aminohapot ovat peräisin hiilihydraattien hajoamisen metabolisista prosesseista;
- skikimiinihappo on sekundaaristen metaboliittien, mutta myös aromaattisten aminohappojen (fenyylialaniini, tryptofaani ja tyrosiini) edeltäjä;
-glykosidi on toissijainen metaboliitti, joka koostuu sokerista ja ei-sokeriyksiköstä, nimeltään aglykoni, joka oletettavasti on peräisin yhdestä yhteenvedosta.
Kaikki biogeeniset rakennuspalikat, joista toissijaiset metaboliitit ovat peräisin, ovat peräisin joko hiilihydraattien kataboliasta tai niiden anaboliasta. Nämä sokerit ovat samat sokeriyksiköt, jotka kerran liittyneet aglykoniin muodostavat glykosideja.
Asetaatin aineenvaihduntareitti on jaettu tiheään biogeneettiseen puuhun, joka sisältää kaikki sen synnyttämien sekundaaristen metaboliittien nimet. Erilaiset, solun tarpeiden mukaan:
- Krebs -sykli ATP: n (ensisijainen aineenvaihdunta) lopputuotannolla;
- rasvahappojen β-hapetus ja synteesi (ensisijainen aineenvaihdunta);
- Synteesi malonaatista tai malonihaposta (4C), joka on peräisin kahden asetaattimolekyylin yhdistymisestä, ja mevalonaatista tai mevalonihaposta (6C), joka on peräisin kolmen asetaattimolekyylin liitosta. Solu käyttää näitä kahta molekyyliä, joissa on parillinen määrä hiiliatomeja, rakentaakseen erilaisia molekyylikategorioita, jotka koostuvat hiilivetyyksiköiden lineaarisista ketjuista, kuten: rasvahapot - joita käytetään vuorostaan glyseridien ja vahojen tuottamiseen - terpenoidit, antrakinonit ja steroidit.
Muut artikkelit aiheesta "Aktiivisten ainesosien biogeneesi ja ominaisuudet"
- Kasvin ensisijainen aineenvaihdunta ja toissijainen aineenvaihdunta
- Farmakognosia
- Skikimiinihapon metaboliareitti